Спинной мозг представляет собой цилиндрической формы вытянутый тяж,
несколько уплощенный спереди назад, расположенный в позвоночном канале. Длина
спинного мозга у мужчин составляет около 45 см, у женщин - 41-42 см. Масса спинного
мозга около 30 г, что составляет 2,3% массы головного мозга. Спинной мозг окружен
тремя оболочками (твердой, паутинной и мягкой). Начинается спинной мозг на уровне
нижнего края большого затылочного отверстия, где переходит в головной мозг.
Нижняя граница суживающегося в виде конуса спинного мозга соответствует
уровню верхнего края второго поясничного позвонка. Ниже этого уровня
находится терминальная нить, окруженная корешками спинномозговых
нервов и оболочками спинного мозга, образующими в нижней части позвоночного
канала замкнутый мешок. В составе терминальной нити различают внутреннюю и наружную
части. Внутренняя часть идет от уровня второго поясничного позвонка до уровня
второго крестцового позвонка, она имеет длину около 15 см. Внутренняя часть
терминальной нити, являющейся остатком конечного отдела эмбрионального спинного
мозга, имеет незначительное количество нервной ткани. Наружная часть терминальной
нити не содержит нервной ткани, является продолжением мозговых оболочек. Она
имеет около 8 см в длину, срастается с надкостницей позвоночного канала на уровне
второго копчикового позвонка (о строении позвоночника см. статью Строение
и функции позвоночника).
Средний диаметр спинного мозга равен 1 см. Спинной мозг имеет два утолщения:
шейное и пояснично-крестцовое, в толще которых располагаются нервные клетки
(о строении нервной ткани см. статью Общее представление
о строении и функциях нервной системы), чьи отростки идут соответственно
к верхним и нижним конечностям. По средней линии на передней поверхности спинного
мозга сверху вниз идет передняя срединная щель. На задней поверхности
ей соответствует менее глубокая задняя срединная борозда. От дна задней
срединной борозды до задней поверхности серого вещества через всю толщину белого
вещества спинного мозга проходит задняя срединная перегородка. На передне-боковой
поверхности спинного мозга, сбоку от передней срединной щели, с каждой стороны
имеется передне-боковая борозда. Через передне-боковую борозду из спинного
мозга выходят передние (двигательные) корешки спинномозговых нервов. На задне-боковой
поверхности спинного мозга с каждой стороны имеется задне-боковая борозда,
через которую в толщу спинного мозга входят нервные волокна (чувствительные)
задних корешков спинномозговых нервов. Эти борозды разделяют белое вещество
каждой половины спинного мозга на три продольных тяжа - канатика: передний,
боковой и задний. Между передней срединной щелью и передне-боковой бороздой
с каждой стороны находится передний канатик спинного мозга. Между
передне-боковой и задне-боковой бороздами на поверхности правой и левой сторон
спинного мозга виден боковой канатик. Позади задне-боковой борозды
по бокам от задней срединной борозды, находится парный задний канатик спинного мозга.
Выходящий через передне-боковую борозду передний корешок образован
аксонами двигательных (моторных) нейронов, залегающих в переднем роге (столбе)
серого вещества спинного мозга. Задний корешок, чувствительный,
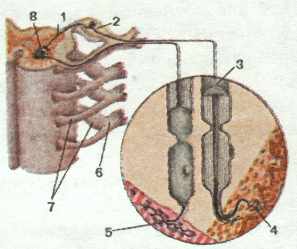
образован совокупностью аксонов псевдоуниполярных нейронов. Тела этих нейронов
образуют спинномозговой узел, располагающийся в позвоночном канале
возле соответствующего межпозвоночного отверстия. В дальнейшем, в межпозвоночном
отверстии, оба корешка соединяются друг с другом, образуя смешанный (содержащий
чувствительные, двигательные и вегетативные нервные волокна) спинномозговой
нерв, который затем делится на переднюю и заднюю ветви. На протяжении спинного
мозга с каждой стороны имеется 31 пара корешков, образующих 31 пару спинномозговых
нервов.
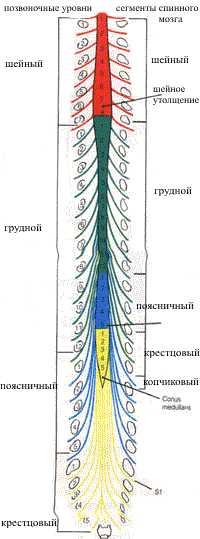
Участок
спинного мозга, соответствующий двум парам корешков спинномозговых нервов (двум
передним и двум задним) называют сегментом спинного мозга. Различают
8 шейных (С1-С8), 12 грудных (Th1-Th12), 5 поясничных (L1-L5), 5 крестцовых
(S1-S5) и 1-3 копчиковых (Co1-Co3) сегмента (всего 31 сегмент). Верхние сегменты
расположены на уровне соотвествующих их порядковому номеру тел шейных позвонков
(рис. 2). Нижние шейные и верхние грудные сегменты находятся на один
позвонок выше, чем тела соответствующих позвонков. В среднем грудном отделе
эта разница равна двум позвонкам, в нижнем грудном - трем позвонкам. Поясничные
сегменты располагаются на уровне тел десятого и одиннадцатого грудных позвонков,
крестцовые и копчиковые сегменты соответствуют уровням двенадцатого грудного
и первого поясничного позвонков. Такое несооветствие сегментов спинного мозга
позвонкам обусловленно разной скоростью роста позвоночника и спинного мозга.
Вначале, на II месяце внутриутробной жизни, спинной мозг занимает весь позвоночный
канал, а затем вследствие более быстрого роста позвоночника отстает в росте
и смещается относительно него вверх. Так что корешки спинномозговых нервов направляются
не только в стороны, но еще и вниз, и тем больше вниз, чем ближе к хвостовому
концу спинного мозга. Направление корешков в поясничной части спинного мозга
внутри позвоночного канала становится почти параллельным продольной оси спинного
мозга, так что мозговой конус и терминальная нить оказываются лежащими среди
густого пучка нервных корешков, который получил название конского хвоста.
В опытах с перерезкой отдельных корешков у животных было установлено, что каждый
сегмент спинного мозга иннервирует три поперечных отрезка, или метамера,
тела: свой собственный, один выше и один ниже. Следовательно, каждый метамер
тела получает чувствительные волокна от трех корешков и, для того чтобы лишить
чувствительности участок тела, необходимо перерезать три корешка (фактор надежности).
Скелетные мышцы (туловища и конечностей) также получают двигательную иннервацию
от трех соседних сегментов спинного мозга. (Более подробно о сегментарном делении
спинного мозга и зонах чувствительной и двигательной иннервации см. в статье Классификация уровня и
степени тяжести травмы спинного мозга American Spinal Injury Association).
Внутреннее строение спинного мозга
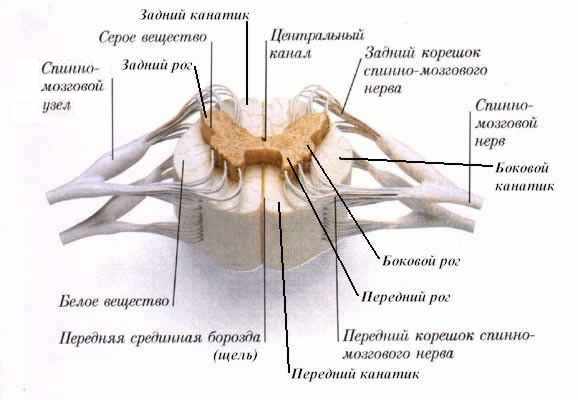
В составе спинного мозга различают серое и белое вещество. Серое вещество располагается
в центральных отделах спинного мозга, белое - на его периферии (рис.1).
Серое вещество спинного мозга
В сером веществе сверху вниз проходит узкий центральный канал.
Вверху канал сообщается с четвертым желудочком головного мозга. Нижний конец
канала расширяется и слепо заканчивается терминальным желудочком (желудочек
Краузе). У взрослого человека местами центральный канал зарастает, его незаросшие
участки содержат спинномозговую жидкость. Стенки канала выстланы эпендимоцитами.
Серое вещество на протяжении спинного мозга с обеих сторон от центрального канала
образует образует два неправильной формы вертикальных тяжа - правый и левый серые столбы. Тонкая пластинка серого вещества, соединяющая спереди от
центрального канала оба серых столба, называется передней серой спайкой.
Сзади от центрального канала правый и левый столбы серого вещества соединены задней серой спайкой. У каждого столба серого вещества выделяют переднюю
часть (передний столб) и заднюю часть (задний столб). На уровне
между восьмым шейным сегментом и вторым поясничным сегментом включительно с
каждой стороны серое вещество образует также латеральное (боковое) выпячивание
- боковой столб. Выше и ниже этого уровня боковые столбы отсутствуют.
На поперечном срезе спинного мозга серое вещество выглядит в виде бабочки или
буквы "Н", а три пары столбов образуют передний, задний и боковой
рога серого вещества. Передний рог более широкий, задний рог -
узкий. Боковой рог топографически соответствует боковому столбу серого
вещества.
Серое вещество спинного мозга образовано телами нейронов, безмиелиновыми и
тонкими миелиновыми волокнами и нейроглией.
В передних рогах (столбах) расположены тела наиболее крупных
нейронов спинного мозга (диаметром 100-140 мкм). Они образуют пять ядер (скоплений). Эти ядра являются моторными (двигательными) центрами спинного мозга.
Аксоны этих клеток составляют основную массу волокон передних корешков спинномозговых
нервов. В составе спинномозговых нервов они идут на периферию и образуют моторные
(двигательные) окончания в мышцах туловища, конечностей и в диафрагме (мышечной
пластине, разделяющей грудную и брюшную полости и играющей главную роль при
вдохе).
Серое вещество задних рогов (столбов) неоднородно. В составе
задних рогов помимо нейроглии имеется большое количество вставочных нейронов,
с которыми контактируют часть аксонов, идущих от чувствительных нейронов в составе
задних корешков. Они представляют собой мелкие мультиполярные, так называемые
ассоциативные и комиссуральные клетки. Ассоциативные нейроны имеют аксоны,
которые заканчиваются на разных уровнях в пределах серого вещества своей половины
спинного мозга. Аксоны комиссуральных нейронов заканчиваются на противоположной
стороне спинного мозга. Отростки нервных клеток заднего рога осуществляют связь
с нейронами выше- и нижележащих соседних сегментов спинного мозга. Отростки
этих нейронов заканчиваются также на нейронах, расположенных в передних рогах
своего сегмента.
В середине заднего рога имеется так называемое собственное ядро. Оно
образовано телами вставочных нейронов. Аксоны этих нервных клеток переходят
в боковой канатик белого вещества (см. ниже) своей и противоположной половины
спинного мозга и участвуют в формировании проводящих путей спинного мозга (переднего
спинно-мозжечкового и спинно-таламического путей).
В основании заднего рога спинного мозга находится грудное ядро (столб
Кларка). Оно состоит из крупных вставочных нейронов (клеток Штиллинга) с хорошо
развитыми, сильно разветвленными дендритами. Аксоны клеток этого ядра входят
в боковой канатик белого вещества своей стороны спинного мозга и также образуют
проводящие пути (задний спинно-мозжечковый путь).
В боковых рогах спинного мозга находятся центры вегетативной
нервной системы. На уровне С8-Th1 расположен симпатический центр расширения
зрачка. В боковых рогах грудного и верхних сегментах поясничного отделов спинного
мозга расположены спинальные центры симпатической нервной системы, иннервирующие
сердце, сосуды, потовые железы, пищеварительный тракт. Именно здесь лежат нейроны,
непосредственно связанные с периферическими симпатическими ганглиями.
Аксоны этих нейронов, образующих вегетативное ядро в сегментах спинного мозга
с восьмого шейного по второй поясничный, проходят через передний рог, выходят
из спинного мозга в составе передних корешков спинномозговых нервов. В крестцовом
отделе спинного мозга заложены парасимпатические центры, иннервирующие
органы малого таза (рефлекторные центры мочеиспускания, дефекации, эрекции,
эякуляции).
Нервные центры спинного мозга являются сегментарными, или рабочими,
центрами. Их нейроны непосредственно связаны с рецепторами и рабочими органами.
Кроме спинного мозга, такие центры имеются в продолговатом и среднем мозге.
Надсегментарные центры, например промежуточного мозга, коры больших полушарий,
непосредственной связи с периферией не имеют. Они управляют ею посредством сегментарных
центров.
Рефлекторная функция спинного мозга.
Серое вещество спинного мозга, задние и передние корешки спинномозговых нервов,
собственные пучки белого вещества образует сегментарный аппарат спинного
мозга. Он обеспечивает рефлекторную (сегментарную) функцию спинного
мозга.
Нервная система функционирует по рефлекторным принципам. Рефлекс представляет
собой ответную реакцию организма на внешнее или внутреннее воздействие и распространяется
по рефлекторной дуге. Рефлекторные дуги - это цепи, состоящие из нервных
клеток.
Простейшая рефлекторная дуга включает чувствительный и эффекторный нейроны,
по которым нервный импульс движется от места возникновения (от рецептора) к
рабочему органу (эффектору) (рис.3). Тело первого чувствительного (псевдоуниполярного)
нейрона находится в спинномозговом узле. Дендрит начинается рецептором, воспринимающим
внешнее или внутреннее раздражение (механическое, химическое и др) и преобразующим
его в нервный импульс, который достигает тела нервной клетки. От тела нейрона
по аксону нервный импульс через чувствительные корешки спинномозговых нервов
направляется в спинной мозг, где образует синапсы с телами эффекторных нейронов.
В каждом межнейронном синапсе с помощью биологически активных веществ (медиаторов)
происходит передача импульса. Аксон эффекторного нейрона выходит из спинного
мозга в составе передних корешков спинномозговых нервов (двигательных или секреторных
нервных волокон) и направляется к рабочему органу, вызывая сокращение мышцы,
усиление (торможение) секреции железы.
Более сложные рефлекторные дуги имеют один или несколько вставочных
нейронов. Тело вставочного нейрона в трехнейронных рефлекторных дугах находится
в сером веществе задних столбов (рогов) спинного мозга и контактирует с приходящим
в составе задних (чувствительных) корешков спинномозговых нервов аксоном чувствительного
нейрона. Аксоны вставочных нейронов направляются к передним столбам (рогам),
где располагаются тела эффекторных клеток. Аксоны эффекторных клеток направляются
к мышцам, железам, влияя на их функцию. В нервной системе много сложных многонейронных
рефлекторных дуг, у которых имеется несколько вставочных нейронов, располагающихся
в сером веществе спинного и головного мозга.
Примером простейшего рефлекса может служить коленный рефлекс, возникающий
в ответ на кратковременное растяжение четырехглавой мышцы бедра легким ударом
по ее сухожилию ниже коленной чашечки. После короткого латентного (скрытого)
периода происходит сокращение четырехглавой мышцы, в результате которого приподнимается
свободно висящая нижняя часть ноги. Коленный рефлекс относится к числу так называемых рефлексов растяжения мышцы, физиологическое значение которых состоит
в регуляции длины мышцы, что особенно важно для поддержания позы. Например,
когда человек стоит, каждое сгибание в коленном суставе, даже такое слабое,
что его невозможно ни увидеть, ни почувствовать, сопровождается растяжением
четырехглавой мышцы и соответствующим усилением активности расположенных в ней
чувствительных окончаний (мышечных веретен). В результате происходит дополнительная
активация мотонейронов четырехглавой мышцы ("коленный рефлекс"), и
повышение ее тонуса, противодействующее сгибанию. И наоборот, слишком сильное
сокращение мышцы ослабляет стимуляцию ее рецепторов растяжения. Частота их импульсации,
возбуждающей мотонейроны, уменьшается, и мышечный тонус ослабевает.
Как правило, в движении участвует несколько мышц, которые по отношению друг
к другу могут выступать как агонисты (действуют в одном направлении)
либо антагонисты (действуют разнонаправленно). Рефлекторный акт возможен
только при сопряженном, так называемом реципрокном торможении двигательных центров мышц-антагонистов. При ходьбе сгибание ноги сопровождается
расслаблением разгибателей и, наоборот, при разгибании тормозятся мышцы-сгибатели.
Если бы этого не происходило, то возникла бы механическая борьба мышц, судороги,
а не приспособительные двигательные акты. При раздражении чувствительного нерва,
вызывающего сгибательный рефлекс, импульсы направляются к центрам мышц-сгибателей
и через специальные вставочные нейроны (тормозные клетки Реншоу) - к
центрам мышц-разгибателей. В первых вызывают процесс возбуждения, а во вторых
- торможения. В ответ возникает координированный, согласованный рефлекторный
акт - сгибательный рефлекс.
Взаимодействие процессов возбуждения и торможения - универсальный принцип,
лежащий в основе деятельности нервной системы. Конечно, он реализуется не только
на уровне сегментов спинного мозга. Вышестоящие отделы нервной системы осуществляют
свое регуляторное влияние, вызывая процессы возбуждения и торможения нейронов
нижестоящих отделов. Важно отметить: чем выше уровень животного, тем сильнее
власть самых высших отделов центральной нервной системы, "тем в большей
степени высший отдел является распорядителем и распределителем деятельности
организма" (И. П. Павлов). У человека таким "распорядителем и распределителем"
является кора больших полушарий головного мозга.
Каждый спинальный рефлекс имеет свое рецептивное поле и свою локализацию (место
нахождения), свой уровень. Так, например, центр коленного рефлекса находится
во II - IV поясничном сегменте; ахиллова - в V поясничном и I - II крестцовых
сегментах; подошвенного - в I - II крестцовом, центр брюшных мышц - в VIII -
XII грудных сегментах. Важнейшим жизненно важным центром спинного мозга является
двигательный центр диафрагмы, расположенный в III - IV шейных сегментах. Повреждение
его ведет к смерти вследствие остановки дыхания.
Кроме двигательных рефлекторных дуг на уровне спинного мозга замыкаются вегетативные
рефлекторные дуги, осуществляющие контроль за деятельностью внутренних органов.
Межсегментарные рефлекторные связи. В спинном мозге помимо описанных
выше рефлекторных дуг, ограниченных пределами одного или нескольких сегментов,
действуют восходящие и нисходящие межсегментарные рефлекторные пути. Вставочными
нейронами в них служат так называемые проприоспинальные нейроны,
тела которых находятся в сером веществе спинного мозга, а аксоны поднимаются
или спускаются на различные расстояния в составе проприоспинальных трактов белого вещества, никогда не покидая спинной мозг. Опыты с дегенерацией нервных
структур (в которых полностью изолируются отдельные части спинного мозга) показали,
что к проприоспинальным нейронам относится большинство его нервных клеток. Некоторые
из них образуют независимые функциональные группы, ответственные за выполнение
автоматических движений (автоматических программ спинного мозга).
Межсегментарные рефлексы и эти программы способствуют координации движений,
запускаемых на разных уровнях спинного мозга, в частности передних и задних
конечностей, конечностей и шеи.
Благодаря этим рефлексам и автоматическим программам спинной мозг способен
обеспечивать сложные согласованные движения в ответ на соответствующий
сигнал с периферии или от вышележащих отделов центральной нервной системы. Здесь
можно говорить о его интегративной (объединяющей) функции спинного мозга,
хотя следует иметь в виду, что у высших позвоночных (в частности, у млекопитающих)
возрастает регуляция спинальных функций высшими отделами центральной нервной
системы (процесс энцефализации).
Спинальная локомоция. Обнаружено, что основные характеристики локомоции,
т. е. перемещения человека или животного в окружающей среде при помощи координированных
движений конечностей, запрограммированы на уровне спинного мозга.
Болевое раздражение какой-либо конечности спинального животного вызывает рефлекторные
движения всех четырех; если же такая стимуляция продолжается достаточно долго,
могут возникнуть ритмичные сгибательные и разгибательные движения не подвергающихся
раздражению конечностей. Если такое животное поставить на тредмилл (бегущую
дорожку), то при некоторых условиях оно будет совершать координированные шагательные
движения, весьма сходные с естественными.
У спинального животного, анестезированного и парализованного кураре, в определенных
условиях можно зарегистрировать ритмично чередующиеся залпы импульсов мотонейронов
разгибателей и сгибателей, примерно соответствующие наблюдаемым при естественной
ходьбе. Поскольку такая импульсация не сопровождается движениями, ее называют ложной локомоцией. Она обеспечивается пока еще не идентифицированными
локомоторными центрами спинного мозга. По-видимому, для каждой конечности существует
один такой центр. Активность центров координируется проприоспинальными системами
и трактами, пересекающими спинной мозг в пределах отдельных ссгменгов.
Предполагают, что у человека тоже есть спинальные локомоторные центры. По-видимому,
их активация при раздражении кожи проявляется в виде шагательного рефлекса
новорожденного. Однако по мере созревания центральной нервной системы
вышестоящие отделы, очевидно, настолько подчиняют себе такие центры. что у взрослого
человека они утрачивают способность к самостоятельной активности. Тем не менее,
активизация локомоторных центров путем интенсивной тренировки лежит в основе
различных методик восстановления ходьбы у больных с повреждением спинного мозга
(см. статью Эффективность
интенсивной тренировки в восстановлении двигательной функции).
Таким образом, даже на уровне спинного мозга обеспечиваются запрограммированные
(автоматические) двигательные акты. Подобные независимые от внешней стимуляции
двигательные программы шире представлены в высших двигательных центрах. Некоторые
из них (например, дыхание) врожденные, другие же (например, езда на велосипеде)
приобретаются в процессе научения.
Белое вещество спинного мозга. Проводниковая функция спинного
мозга.
Белое вещество спинного мозга образовано совокупностью продольно ориентированных
нервных волокон, идущих в восходящем или нисходящем направлении. Белое вещество
окружает со всех сторон серое и разделяется, как уже упомянуто было выше, на
три канатика: передний, задний, боковой.
Кроме этого в нем выделяют переднюю белую спайку. Она располагается
кзади от передней срединной щели и соединяет передние канатики правой и левой
сторон.
Пучки нервных волокон (совокупность отростков) в канатиках спинного мозга составляют проводящие пути спинного мозга. Различают три системы пучков:
Короткие пучки ассоциативных волокон связывают сегменты спинного
мозга, расположенные на различных уровнях.
Восходящие (афферентные, чувствительные) пути направляются
к центрам головного мозга.
Нисходящие (эфферентные, двигательные) пути идут от головного
мозга к клеткам передних рогов спинного мозга.
В белом веществе передних канатиков проходят в основном нисходящие проводящие
пути, в боковых канатиках - восходящие и нисходящие, в задних канатиках - восходящие
проводящие пути.
Чувствительные (восходящие) пути. Спинной мозг проводит четыре вида
чувствительности: тактильную (чувство прикосновения и давления), температурную,
болевую и проприоцептивную (от рецепторов мышц и сухожилий, так называемое суставно-мышечное
чувство, чувство положения и движения тела и конечностей).
Основная масса восходящих путей проводит проприоцептивную чувствительность. Это говорит о важности контроля движений, так называемой обратной связи, для
двигательной функции организма. Пути проприоцептивной чувствительности направляются
к коре полушарий большого мозга и в мозжечок, который участвует в координации
движений. Проприоцептивный путь к коре больших полушарий представлен двумя пучками:
тонким и клиновидным. Тонкий пучок (пучок Голля) проводит импульсы
от проприорецепторов нижних конечностей и нижней половины тела и прилежит к
задней срединной борозде в заднем канатике. Клиновидный пучок (пучок Бурдаха) примыкает к нему снаружи и несет импульсы от верхней половины туловища и от
верхних конечностей. К мозжечку идут два спинно-мозжечковых пути - передний (Флексига) и задний (Говерса). Они располагаются в составе боковых
канатиков. Передний спинно-мозжечковый путь служит для контроля положения конечностей
и равновесия всего тела во время движения и позы. Задний спинно-мозжечковый
путь специализирован для быстрой регуляции тонких движений верхних и нижних
конечностей. Благодаря поступлению импульсов от проприоцепторов мозжечок участвует
в автоматической рефлекторной координации движений. Особенно отчетливо это проявляется
при внезапных нарушениях равновесия во время ходьбы, когда в ответ на изменение
положения тела возникает целый комплекс непроизвольных движений, направленный
на поддержание равновесия.
Импульсы болевой и температурной чувствительности проводит латеральный (боковой) спинно-таламический путь. Первым
нейроном этого пути являются чувствительные клетки спинномозговых узлов. Их
периферические отростки (дендриты) приходят в составе спинномозговых нервов.
Центральные отростки образуют задние корешки и идут в спинной мозг, оканчиваясь
на вставочных нейронах задних рогов (2-й нейрон). Отростки вторых нейронов через
переднюю белую спайку переходят на противоположную сторону (образуют перекрест)
и поднимаются в составе бокового канатика спинного мозга в головной мозг. В
результате того, что волокна по пути перекрещиваются, импульсы от левой половины
туловища и конечностей передаются в правое полушарие, а от правой половины -
в левое.
Тактильную чувствительность (чувство осязания, прикосновения, давления) проводит передний спинно-таламический путь, идущий в составе переднего
канатика спинного мозга.
Двигательные пути представлены двумя группами:
1. Передний и боковой (латеральный) пирамидные (кортико-спинальные) пути,
проводящие импульсы от коры к двигательным клеткам спинного мозга, являющиеся
путями произвольных (осознанных) движений. Они представлены аксонами гигантских
пирамидных клеток (клеток Беца), залегающих в коре предцентральной извилины
полушарий большого мозга. На границе со спинным мозгом большая часть волокон
общего пирамидного пути переходит на противоположную сторону (образует перекрест)
и образует боковой пирамидный путь, который спускается в боковом канатике
спинного мозга, заканчиваясь на мотонейронах переднего рога. Меньшая часть волокон
не перекрещивается и идет в переднем канатике, образуя передний пирамидный
путь. Однако и эти волокна также постепенно переходят через переднюю белую
спайку на противоположную сторону (образуют посегментный перекрест) и заканчиваются
на двигательных клетках переднего рога. Отростки клеток переднего рога образуют
передний (двигательный) корешок и заканчиваются в мышце двигательным окончанием.
Таким образом, оба пирамидных пути являются перекрещенными. Поэтому при одностороннем
повреждении головного или спинного мозга возникают двигательные нарушения ниже
места повреждения на противоположной стороне тела. Пирамидные пути - двухнейронные
(центральный нейрон - пирамидная клетка коры, периферический нейрон - мотонейрон переднего рога спинного мозга). При повреждении тела или аксона
центрального нейрона наступает центральный (спастический) паралич,
а при повреждении тела или аксона периферического нейрона - периферический
(вялый) паралич.
2. Экстрапирамидные, рефлекторные двигательные пути. К ним относятся:
- красноядерно-спинномозговой (руброспинальный) путь - идет в составе
боковых канатиков от клеток красного ядра среднего мозга к передним рогам спинного
мозга, несет импульсы подсознательного управления движениями и тонусом скелетных
мышц;
- текто-спинальный (покрышечно-спинальный) путь - идет в переднем канатике,
связывает верхние холмики покрышки среднего мозга (подкорковые центры зрения)
и нижние холмики (центры слуха) с двигательными ядрами передних рогов спинного
мозга, функция его заключается в обеспечении координированных движений глаз,
головы и верхних конечностей на неожиданные световые и звуковые воздействия;
- вестибуло-спинальный (предверно-спинальный) путь - направляется от
преддверных (вестибулярных) ядер (8-й пары черепных нервов) к двигательным клеткам
передних рогов спинного мозга, оказывает возбуждающее влияние на двигательные
ядра мышц-разгибателей (антигравитационная мускулатура), причем преимущественно
на осевые мышцы (мышцы позвоночного столба) и на мышцы поясов верхних и нижних
конечностей. На сгибательную мускулатуру вестибуло-спинальный тракт оказывает
тормозящее влияние.
Кровоснабжение спинного мозга.
Спинной мозг кровоснабжается продольно идущими передней и двумя задними спинномозговыми
артериями. Передняя спинномозговая артерия образуется при соединении спинномозговых
ветвей правой и левой позвоночных артерий, и идет вдоль передней продольной
щели спинного мозга. Задняя спинномозговая артерия, парная, прилежит к задней
поверхности спинного мозга возле вхождения в него заднего корешка спинномозгового
нерва. Эти артерии продолжаются на протяжении всего спинного мозга. Они соединяются
со спинномозговыми ветвями глубокой шейной артерии, задних межреберных, поясничных
и латеральных крестцовых артерий, проникающими в позвоночный канал через межпозвоночные
отверстия.
Вены спинного мозга впадают во внутреннее позвоночное венозное сплетение.
Снаружи располагается твердая мозговая оболочка. Между этой оболочкой
и надкостницей позвоночного канала находится эпидуральное пространство.
Кнутри от твердой мозговой оболочки имеется паутинная оболочка,
отделенная от твердой мозговой оболочки субдуральным пространством. Непосредственно
к спинному мозгу прилежит внутренняя мягкая мозговая оболочка.
Между паутинной и внутренней мозговой оболочками располагается подпаутинное
(субарахноидальное) пространство, заполненное спинномозговой жидкостью.
Твердая оболочка спинного мозга представляет собой слепой мешок, внутри
которого находятся спинной мозг, передние и задние корешки спинномозговых нервов
и остальные мозговые оболочки. Твердая мозговая оболочка плотная, образована
волокнистой соединительной тканью, содержит значительное количество эластических
волокон. Вверху твердая оболочка спинного мозга прочно срастается с краями большого
затылочного отверстия и переходит в твердую оболочку головного мозга. В позвоночном
канале твердая мозговая оболочка укрепляется ее отростками, продолжающимися
в оболочки спинномозговых нервов. Эти отростки срастаются с надкостницей в области
межпозвоночных отверстий. Твердую мозговую оболочку укрепляют также многочисленные
фиброзные пучки, идущие к задней продольной связке позвоночника. Эти пучки лучше
выражены в шейной, поясничной и крестцовой областях и хуже - в грудной области.
В верхнем шейном отделе твердая оболочка покрывает правую и левую позвоночные
артерии.
Наружная поверхность твердой мозговой оболочки отделена от надкостницы эпидуральным
пространством. Оно заполнено жировой клетчаткой и содержит внутреннее
позвоночное венозное сплетение. Внутренняя поверхность твердой оболочки спинного
мозга отделена от паутинной оболочки щелевидным субдуральным пространством.
Оно заполнено большим количеством тонких соединительнотканных пучков. Субдуральное
пространство спинного мозга вверху сообщается с одноименным пространством головного
мозга, внизу слепо заканчивается на уровне второго крестцового позвонка. Ниже
этого уровня пучки фиброзных волокон твердой мозговой оболочки продолжаются
в терминальную нить.
Паутинная оболочка спинного мозга представлена тонкой полупрозрачной
соединительнотканной пластинкой, расположенной кнутри от твердой оболочки. Твердая
и паутинная оболочки срастаются между собой только возле межпозвоночных отверстий.
Между паутинной и мягкой оболочками (в субарахноидальном пространстве) расположена
сеть перекладин, состоящих из тонких пучков коллагеновых и эластических волокон.
Эти соединительнотканные пучки соединяют паутинную оболочку с мягкой оболочкой
и со спинным мозгом.
Мягкая (сосудистая) оболочка спинного мозга плотно прилежит в поверхности
спинного мозга. Соединительнотканные волокна, отходящие от мягкой оболочки,
сопровождают кровеносные сосуды, заходят вместе с ними в ткань спинного мозга.
Между паутинной и мягкой мозговыми оболочками находится подпаутинное,
или субарахноидальное пространство. В нем содержится 120-140 мл
спинномозговой жидкости. В верхних отделах это пространство продолжается в подпаутинное
пространство головного мозга. В нижних отделах подпаутинное пространство спинного
мозга содержит лишь корешки спинномозговых нервов. Ниже уровня второго поясничного
позвонка пунктированием возможно получить для исследования спинномозговую жидкосгь,
не рискуя повредить спинной мозг.
От боковых сторон мягкой мозговой оболочки спинного мозга, между передними и
задними корешками спинномозговых нервов вправо и влево фронтально идет зубчатая
связка. Зубчатая связка также срастается с паутинной и с внутренней
поверхностью твердой оболочки спинного мозга, связка как бы подвешивает спинной
мозг в субарахноидальном пространстве. Имея сплошное начало на боковых поверхностях
спинного мозга, связка в латеральном направлении разделяется на 20-30 зубцов.
Верхний зубец соответствует уровню большого затылочного отверстия, нижний расположен
между корешками двенадцатого грудного и первого поясничного позвонков. Помимо
зубчатых связок спинной мозг фиксируется в позвоночном канале при помощи задней
подпаутинной перегородки. Эта перегородка начинается от твердой, паутинной и
мягкой оболочек и соединяется с задней срединной перегородкой, имеющейся между
задними канатиками белого вещества спинного мозга. В нижней поясничной и крестцовой
областях спинного мозга задняя перегородка подпаутинного пространства, как и
зубчатые связки, отсутствует. Жировая клетчатка и венозные сплетения эпидурального
пространства, оболочки спинного мозга, спинномозговая жидкость и связочный аппарат
предохраняют спинной мозг от сотрясений при движениях тела.
Литература
1. Антонен Е.Г. Спинной мозг (анатомо-физиологические и неврологические аспекты).
- Учебное пособие. На сайте .
2. Сапин М.Р., Никитюк Д.Б. Анатомия человека. - В 3 томах. - М. - 1998. - Т.3.
3. Материалы сайта .
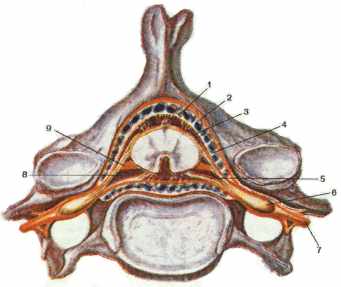
 Участок
спинного мозга, соответствующий двум парам корешков спинномозговых нервов (двум
передним и двум задним) называют сегментом спинного мозга. Различают
8 шейных (С1-С8), 12 грудных (Th1-Th12), 5 поясничных (L1-L5), 5 крестцовых
(S1-S5) и 1-3 копчиковых (Co1-Co3) сегмента (всего 31 сегмент). Верхние сегменты
расположены на уровне соотвествующих их порядковому номеру тел шейных позвонков
(рис. 2). Нижние шейные и верхние грудные сегменты находятся на один
позвонок выше, чем тела соответствующих позвонков. В среднем грудном отделе
эта разница равна двум позвонкам, в нижнем грудном - трем позвонкам. Поясничные
сегменты располагаются на уровне тел десятого и одиннадцатого грудных позвонков,
крестцовые и копчиковые сегменты соответствуют уровням двенадцатого грудного
и первого поясничного позвонков. Такое несооветствие сегментов спинного мозга
позвонкам обусловленно разной скоростью роста позвоночника и спинного мозга.
Вначале, на II месяце внутриутробной жизни, спинной мозг занимает весь позвоночный
канал, а затем вследствие более быстрого роста позвоночника отстает в росте
и смещается относительно него вверх. Так что корешки спинномозговых нервов направляются
не только в стороны, но еще и вниз, и тем больше вниз, чем ближе к хвостовому
концу спинного мозга. Направление корешков в поясничной части спинного мозга
внутри позвоночного канала становится почти параллельным продольной оси спинного
мозга, так что мозговой конус и терминальная нить оказываются лежащими среди
густого пучка нервных корешков, который получил название конского хвоста.
Участок
спинного мозга, соответствующий двум парам корешков спинномозговых нервов (двум
передним и двум задним) называют сегментом спинного мозга. Различают
8 шейных (С1-С8), 12 грудных (Th1-Th12), 5 поясничных (L1-L5), 5 крестцовых
(S1-S5) и 1-3 копчиковых (Co1-Co3) сегмента (всего 31 сегмент). Верхние сегменты
расположены на уровне соотвествующих их порядковому номеру тел шейных позвонков
(рис. 2). Нижние шейные и верхние грудные сегменты находятся на один
позвонок выше, чем тела соответствующих позвонков. В среднем грудном отделе
эта разница равна двум позвонкам, в нижнем грудном - трем позвонкам. Поясничные
сегменты располагаются на уровне тел десятого и одиннадцатого грудных позвонков,
крестцовые и копчиковые сегменты соответствуют уровням двенадцатого грудного
и первого поясничного позвонков. Такое несооветствие сегментов спинного мозга
позвонкам обусловленно разной скоростью роста позвоночника и спинного мозга.
Вначале, на II месяце внутриутробной жизни, спинной мозг занимает весь позвоночный
канал, а затем вследствие более быстрого роста позвоночника отстает в росте
и смещается относительно него вверх. Так что корешки спинномозговых нервов направляются
не только в стороны, но еще и вниз, и тем больше вниз, чем ближе к хвостовому
концу спинного мозга. Направление корешков в поясничной части спинного мозга
внутри позвоночного канала становится почти параллельным продольной оси спинного
мозга, так что мозговой конус и терминальная нить оказываются лежащими среди
густого пучка нервных корешков, который получил название конского хвоста.